
انعطافپذیری تکاملی برای کسب یا از دست دادن پیچیدگی دندان، جرقهای برای تنوعزایی ماهیها
چرا احتمال تنوعزایی برخی گونهها بیشتر از سایرین است؟ برای بزرگترین گروه مهرهداران، یعنی ماهیان پرتو باله، توانایی تکامل با انتقال بین دندانهای ساده و پیچیده، به تنوعزایی آنها دامن زده است.
تنوع زیستی شگفتانگیز است، اما همه گونهها برای تشکیل گونههای جدید متنوع نمیشوند - فرآیندی که به عنوان گونهزایی شناخته میشود. بنابراین، چرا برخی از جمعیتها تقسیم میشوند و برخی دیگر نه؟ در Nature، Peoples و همکاران.1 گزارش تحقیق خود را در مورد تنوع ماهیها از کلاس Actinopterygii (ماهیان پرتو باله)، که بزرگترین گروه مهرهداران هستند و بیش از 35000 گونه زنده را شامل میشوند، ارائه میدهند.
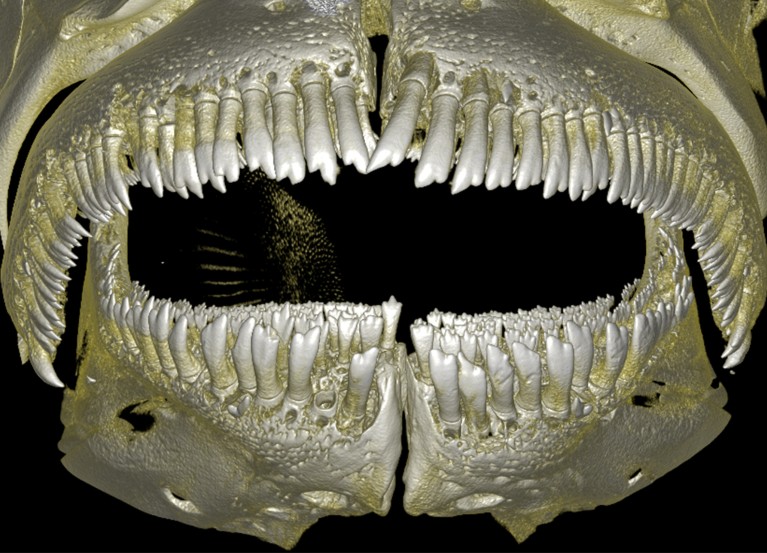
نویسندگان دندانها را به دو گروه برای بیش از 88 درصد از گونههای زنده ماهیان پرتو باله طبقهبندی کردند. Peoples و همکاران یک مجموعه داده جامع از این که آیا دندانها ساده (شکل 1) با یک پشته (کاسپ) هستند، که در مورد دندان جلویی انسان صادق است، یا پیچیده (شکل 2) با بیش از یک کاسپ، که در مورد دندانهای آسیاب انسان صادق است، گردآوری کردند. شواهد نشان میدهد که موفقیت تکاملی این ماهیها لزوماً ناشی از نوآوری کلیدی دندانهای پیچیده نبوده، بلکه ناشی از انعطافپذیری در به دست آوردن و از دست دادن پیچیدگی دندان (ناپایداری پیچیدگی دندان) بوده است.
ممکن است منابع فراوانی به راحتی در محیط زیست در دسترس باشند، اما موجودات زنده میتوانند فاقد ویژگیهای (صفاتی) مورد نیاز برای بهرهبرداری از آنها باشند. راهی برای پر کردن این شکاف و فراهم کردن دسترسی گونهها به منابع یا فرصتهای بومشناختی که قبلاً غیرقابل دسترس بودند، میتواند زمانی ایجاد شود که یک صفت جدید تکامل یابد، که به آن نوآوری کلیدی2 گفته میشود. بنابراین، تصور میشود که این نوآوریها بخش عمدهای از تنوع روی زمین را ایجاد کردهاند3. به عنوان مثال، تکامل کاسپهای متعدد روی دندانها، که به عنوان پیچیدگی دندان شناخته میشود، گزینههای رژیم غذایی را گسترش داد و به تنوعزایی در مهرهداران دامن زد.
Peoples و همکاران. عمیقتر به این رابطه بین پیچیدگی دندان و تنوعزایی پرداختند. آنها نشان میدهند که ناپایداری به شدت با تنوعزایی مرتبط است، به طوری که ناپایداری بالا به شدت با تنوعزایی بیشتر مرتبط است. در ماهیان پرتو باله، بیشتر دودمانهایی که ناپایداری تکاملی کمی داشتند، تنوع کمی نیز داشتند. یک استثنا در این تنوع کم ماهیان پرتو باله، برای ماهیان سیچلاید آفریقایی، Liparidae، Mormyridae و Serrasalmidae بود که تحت سلطه دودمانهایی بودند که پیچیدگی دندان بالایی از خود نشان میدادند، و همچنین نرخهای بالای گونهزایی و نرخهای بالای تکامل اشکال جدید. سیچلایدهای آفریقایی، همچنین به عنوان سیچلیدها شناخته میشوند، متنوعترین و مثال کتاب درسی از تنوع گونهها برای پر کردن جایگاههای مختلف (پدیدهای که به عنوان تابش تطبیقی شناخته میشود)، تحت سلطه دودمانهایی بودند که نه تنها به سرعت بین دندانهای ساده و پیچیده (ناپایداری بالا) انتقال مییافتند، بلکه نسبت بالایی از دندانهای پیچیده در گونههای فردی معین داشتند.
سیچلایدهای آفریقایی به دلیل تابشهای دیدنی خود شناخته شدهاند. تقریباً 2000 گونه منحصر به فرد در دریاچههای آفریقای شرقی تکامل یافتهاند، به طوری که دریاچههای بزرگ مانند دریاچه تانگانیکا 250 گونه، دریاچه مالاوی 700 گونه و دریاچه ویکتوریا 700 گونه بومی دارند4. این تنوع غنی همواره محققانی را که در مورد دلیل تکامل آن تحقیق میکنند، جذب کرده است. موضوع تکراری در مطالعات قبلی نشان میدهد که تنوع گونهای استثنایی ناشی از فرصتهای بومشناختی فوقالعاده5، و ترکیبی از نوآوریهای کلیدی و پتانسیل ژنومی6,7 بوده است. مطالعه Peoples و همکاران تکرار میکند که سیچلایدهای آفریقایی با انتقال سریع بین دندانهای ساده و پیچیده از فرصتهای بومشناختی رو به رشد استفاده کردند، و این امر واگرایی همزمان در رژیم غذایی و در امتداد زیستگاههایی را تسهیل کرد که باعث تنوعزایی سریع شد.
هنگامی که نویسندگان مجموعه داده پیچیدگی دندان را گسترش دادند تا تمام جنسهای سیچلاید را نشان دهند، که 92.7٪ از گونههای سیچلاید توصیف شده را پوشش میداد، دریافتند که سیچلایدهای آفریقایی در مقایسه با سایر اکتینوپتریژیانها نرخهای بینظیری از تنوعزایی دارند. نسبت زیادی از سیچلایدها دندانهای پیچیده داشتند و سیچلایدها نیز ناپایداری بالایی در پیچیدگی دندان داشتند. به نظر میرسد این ترکیب باعث پیشرفت تنوعزایی سیچلایدها شده است، و نویسندگان نتیجه میگیرند که ناپایداری پیچیدگی دندان نشان داده شده در سیچلایدهای آفریقایی خود یک نوآوری کلیدی است. راههای متعددی وجود دارد که میتوان یک صفت را به عنوان یک نوآوری کلیدی طبقهبندی کرد، اما به طور کلی، باید شواهدی از افزایش تنوعزایی پس از تکامل صفت وجود داشته باشد. این در برخی از مطالعات نشان داده شده است - یک مثال رایج، گیاهان گلدار به نام آنژیوسپرم است، که در آن افزایش خیرهکنندهای در تنوع گونهها زمانی رخ داد که ساختارهایی به نام مهمیزهای شهد گل تکامل یافتند8.
مطالعه Peoples و همکاران به درک دانشمندان از تکامل نوآوریهای کلیدی و ناپایداری پیچیدگی دندان که منجر به تنوعزایی شده است، میافزاید. نتایج آنها دیدگاه دیگری را در مورد اینکه چرا ماهیان پرتو باله گونههای بسیار زیادی دارند و مهمتر از همه، چه چیزی تنوعزایی سیچلایدهای آفریقایی را ممکن ساخته است، ارائه میدهد.
این مطالعه یک مسیر جدید تحقیقاتی را برای بررسی ناپایداری پیچیدگی دندان به عنوان یک نوآوری کلیدی در سیچلایدهای آفریقایی نشان میدهد، و مطالعات آینده از جفت کردن این با رویکردهای دیگر، مانند تجزیه و تحلیل ژنومی، سود خواهند برد. شواهد نشان میدهد که تجزیه و تحلیل کل ژنوم بینش اساسی در مورد مولفهای از تکامل نوآوریهای کلیدی ارائه میدهد. به عنوان مثال، یکی از نمونههای رایج ذکر شده از یک نوآوری کلیدی به نام حلق و دندانی - تغییراتی در آروارههای حلقی که در برخی از کلادهای بسیار غنی از گونهها، مانند سیچلایدها یافت میشود - دوباره ارزیابی شده است زیرا تجزیه و تحلیل ژنومی هیچ شواهدی از افزایش تنوعزایی مرتبط با آن نشان نمیدهد9. این نشان میدهد که در آینده، ادغام رویکردهای متعدد یک تاکتیک قدرتمند خواهد بود و بینشهای جدیدی را برای مطالعات نوآوریهای کلیدی اضافه خواهد کرد.
منابع
-
Peoples, N., Burns, M. D., Mihalitsis, M. & Wainwright, P. C. Nature https://doi.org/10.1038/s41586-025-08612-z (2025).
-
Cracraft, J. in Evolutionary Innovations (ed. Nitecki, M. H.) (Univ. Chicago Press, 1990).
-
Miller, A. H., Stroud, J. T. & Losos, J. B. Trends Ecol. Evol. 38, 122–131 (2023).
-
Turner, G. F., Seehausen, O., Knight, M. E., Allender, C. J. & Robinson, R. L. Mol. Ecol. 10, 793–806 (2001).
-
Losos, J. B. & Mahler, D. L. in Evolution Since Darwin (eds Bell, M. A. et al.) 381–420 (Oxford Univ. Press, 2010).
-
Meier, J. I. et al. Nature Commun. 8, 14363 (2017).
-
Meier, J. I. et al. Nature. Commun. 10, 5391 (2019).
-
Hodges, S. A. & ; Stpiczynska, M. New Phytol. 230, 488–502 (2021).
-
Muschick, M., Roesti, M., Indermaur, A., Matthews, B. & Salzburger, W. Ecol. Lett. 15, 190–200 (2012).