چکیده
آسیبهای روحی مادرانه بر نتایج سلامتی نوزاد و بزرگسال تأثیر میگذارند و ممکن است از طریق تغییرات اپیژنتیکی مانند متیلاسیون DNA (DNAm) بر نسلهای آینده نیز اثر بگذارند. تحقیقات در مورد انتقال بین نسلی اپیژنتیکی اثرات ناشی از آسیبهای روحی در انسان محدود است. در این مطالعه، ما نشانههای DNAm خشونتهای مرتبط با جنگ را با مقایسه مواجهههای سلول جنسی، پیش از تولد و مستقیم با خشونت در بین سه نسل از پناهندگان سوری ارزیابی کردیم. ما خانوادههایی را که در آنها مادربزرگ باردار در مقابل مادر باردار در معرض خشونت قرار گرفته بودند، مقایسه کردیم و یک گروه کنترل بدون مواجهه با جنگ را نیز در نظر گرفتیم. ما نمونههای سواب دهانی و دادههای نظرسنجی را از مادران و ۱ تا ۲ فرزند در هر یک از ۴۸ خانواده (تعداد شرکتکنندگان=۱۳۱ نفر) جمعآوری کردیم. بر اساس یک مطالعه انجمنی در مورد کل ژنوم (EWAS)، مناطق متیله شده متفاوتی (DMPs) را شناسایی کردیم: ۱۴ مورد با سلول جنسی و ۲۱ مورد با مواجهه مستقیم با خشونت مرتبط بودند. بیشتر DMPsها جهت یکسانی را در تغییر DNAm در سراسر مواجهههای سلول جنسی، پیش از تولد و مستقیم نشان دادند که نشاندهنده یک پاسخ اپیژنتیکی مشترک به خشونت است. علاوه بر این، ما تسریع سن اپیژنتیکی را در ارتباط با مواجهه پیش از تولد با خشونت در کودکان شناسایی کردیم که بر دوره بحرانی رشد درون رحم تأکید دارد. این اولین گزارش از یک نشانه اپیژنتیکی بین نسلی خشونت است که پیامدهای مهمی برای درک وراثت آسیبهای روحی دارد.
مقدمه
آسیبهای روحی مادرانه بر سلامت جنین در دوران بارداری و در طول بزرگسالی تأثیر میگذارد و ممکن است از طریق تغییرات اپیژنتیکی مانند متیلاسیون DNA (DNAm) 1,2,3,4 بر نسلهای آینده نیز اثر بگذارد. استرسورهای مادرانه که تأثیر شناختهشدهای بر جنین در حال رشد دارند عبارتند از کمبودهای تغذیهای 5 و قرار گرفتن در معرض سموم 6 و همچنین استرسورهای روانی-اجتماعی مانند اضطراب 7 و، به طور بالقوه، قرار گرفتن در معرض خشونت و آسیبهای روحی 8,9. فرضیه منشأهای رشدی سلامت و بیماری (DOHaD) تأثیر ناگواریهای اولیه زندگی بر نتایج سلامتی در دوران بعدی زندگی را با شناسایی ارتباطات قوی بین ناگواریهای اولیه زندگی، مانند وزن کم هنگام تولد و شرایط نامساعد زندگی، با افزایش خطر ابتلا به بیماریهای قلبی عروقی در بزرگسالی رسمیت بخشید 10,11. طبق اصول DOHaD، جنین در حال رشد با انعطافپذیری بالای فنوتیپی مشخص میشود و از نشانههای محیطی برای تعیین یک فنوتیپ بهینه برای زنده ماندن در محیط پس از تولد استفاده میکند. با این حال، انطباق بر اساس نشانههای داخل رحمی ممکن است فنوتیپی را انتخاب کند که در نهایت در دوران بعدی زندگی ناسازگار باشد، بنابراین خطر ابتلا به برخی بیماریها را افزایش میدهد 3,12,13.
تصور میشود که اثرات استرسورهای روانی-اجتماعی و آسیبهای روحی از طریق تغییرات سلولی در محور هیپوتالاموس-هیپوفیز-آدرنال (HPA) مادر و جنین و تغییرات همراه در متابولیسم گلوکوکورتیکوئید 3 از مادر به فرزند منتقل میشود. با این حال، مکانیسم مولکولی اساسی که اطلاعات مربوط به آسیبهای روحی را برای دههها در بزرگسالی رمزگذاری و حفظ میکند، به خوبی درک نشده است. با گسترش قدرت پیشبینیکننده فرضیه DOHaD، مکانیسمهای اپیژنتیکی پیشنهاد شدهاند که تأثیر استرسهای روانی-اجتماعی و آسیبهای روحی را بر نسلهای آینده تعدیل میکنند 14,15. به طور خاص، تغییرات اپیژنتیکی حساس به محیط ممکن است در مکانهای انتخابی در اپیژنوم انسان تکامل یافته باشند تا در مقایسه با ژنوم که با سرعت کمتری تکامل مییابد، یک پاسخ سریع و کوتاهمدت به استرسورهای محیطی ارائه دهند. علاوه بر این، زیرمجموعهای از آن تغییرات اپیژنتیکی ممکن است تکامل یافته باشند تا قابل وراثت باشند و مزایای انتخابی حساسیت محیطی را به نسلهای آینده منتقل کنند. طیف وسیعی از تغییرات اپیژنتیکی وجود دارد، از جمله متیلاسیون DNA (DNAm)، تغییر هیستون و RNAهای غیرکدکننده، که به تنظیم بیان ژن کمک میکنند. DNAm معمولاً به افزودن یک گروه متیل به یک باز نوکلئوتیدی سیتوزین اشاره دارد که به دنبال آن یک باز گوانین (یک سایت CpG) میآید. تغییرات در DNAm بسته به ژن و زمینه ژنوم و همچنین مرحله رشد و فعالیت تکثیر ژنوم 16 میتواند منجر به افزایش یا کاهش در بیان ژن شود. DNAm به خوبی مورد مطالعه قرار گرفته است زیرا یکی از عوامل اصلی اپیژنتیکی است که تنظیم ژن را در پستانداران کنترل میکند، نقش مهمی در تمایز و برنامهریزی مجدد سلولی ایفا میکند و به تغییرات محیطی حساس است 17.
شواهد قابل توجهی از این فرضیه پشتیبانی میکنند که تغییرات در DNAm ممکن است تأثیر استرس و آسیبهای روحی مادرانه را بر طیفی از نتایج سلامتی فرزندان تعدیل کند. به طور خاص، استرس و آسیبهای روحی مادرانه با تغییرات در DNAm نوزاد و تسریع سن اپیژنتیکی و همچنین شاخصهای بدتر شدن نتایج سلامتی مانند دیابت 18,19,20,21,22 مرتبط بودهاند. این ایده که نشانههای DNAm مرتبط با آسیبهای روحی ممکن است به نسلهای آینده در انسان منتقل شوند، گمانهزنی بیشتری است 4,23,24. انتقال به نسلهای F2 و F3 به عنوان وراثت اپیژنتیکی بین نسلی در نظر گرفته میشود، در حالی که انتقال به نسل F4 غیرمواجهه و فراتر از آن، وراثت اپیژنتیکی فرا نسلی نامیده میشود 14. اگرچه اپیژنوم پستانداران در طول گامتوژنز و جنینزایی برای امکان برنامهریزی مجدد اپیژنتیکی از بیشتر نشانههای DNAm پاک میشود 25، اما حمایت جالبتوجهی از مدلهای حیوانی برای نشانههای اپیژنتیکی ناشی از محیط وجود دارد که در برابر برنامهریزی مجدد اپیژنتیکی مقاومت میکنند و به صورت بین نسلی و فرا نسلی با اثرات فنوتیپی به ارث میرسند 26,27. وراثت اپیژنتیکی فرا نسلی، از جمله در پاسخ به محرکهای محیطی، به خوبی در مخمر، گیاهان، ماهی، Caenorhabditis elegans و Drosophila مستند شده است 28,29,30,31,32,33. مثالهای کمتری از وراثت اپیژنتیکی در مدلهای حیوانی وجود دارد، اما یک مطالعه اخیر وراثت DNAm را در موشها تا چهار نسل گزارش کرده است 34. در مقابل، تنها یک مطالعه در انسان ارتباط بین قرار گرفتن مادربزرگها در معرض آسیبهای روحی و تغییرات DNAm در نوهها را گزارش کرده است 35. مطالعه وراثت اپیژنتیکی در انسان به دلیل دشواری انجام مطالعات تجربی چند نسلی و همچنین اهمیت عوامل فرهنگی و محیطی 4 پیچیده است. به طور کلی، تغییرات در DNAm در انسان پس از لقاح پیچیده و به طور پیچیدهای زمانبندی شده است که گواهی بر اهمیت DNAm در اوایل رشد 36 و حمایت از این احتمال است که تعداد کمی از نشانههای DNAm ممکن است تکامل یافته باشند که به محیط حساس هستند و به نسلهای آینده قابل انتقال هستند.
در این مطالعه، ما از فرضیه DOHaD به عنوان چارچوبی برای بررسی تغییرات اپیژنتیکی به عنوان مکانیزمی استفاده میکنیم که تأثیر آسیبهای روانی-اجتماعی را بر نسلهای آینده تعدیل میکند. به طور خاص، ما وجود موقعیتهای متیله شده متفاوتی (DMPs) را در اپیژنوم انسان پیشنهاد میکنیم که به محیط روانی-اجتماعی، به ویژه خشونت، حساس هستند و به نسلهای آینده قابل انتقال هستند. ما نتایج حاصله از یک مطالعه انجمنی در مورد کل ژنوم (EWAS) را ارائه میدهیم که در آن ارتباط بین DNAm و خشونتهای مرتبط با جنگ را در یک گروه از خانوادههای پناهنده سوری با مواجهههای مستقیم، پیش از تولد و سلول جنسی متضاد با خشونت جنگ آزمایش میکنیم. گروه مطالعه سه نسلی ما با مواجهههای متمایز با خشونت، در میان مطالعات انسانی در تمرکز مستقیم بر نشانههای اپیژنتیکی بین نسلی جدید است. فرضیه ما، یعنی اینکه قرار گرفتن در معرض خشونت ممکن است نشانههای اپیژنتیکی بین نسلی ایجاد کند، پیامدهای عمیقی برای درک ما از تکامل دارد، مانند اینکه چگونه یک تجربه زیسته میتواند به صورت بین نسلی در ژنوم جاسازی شود.
نتایج
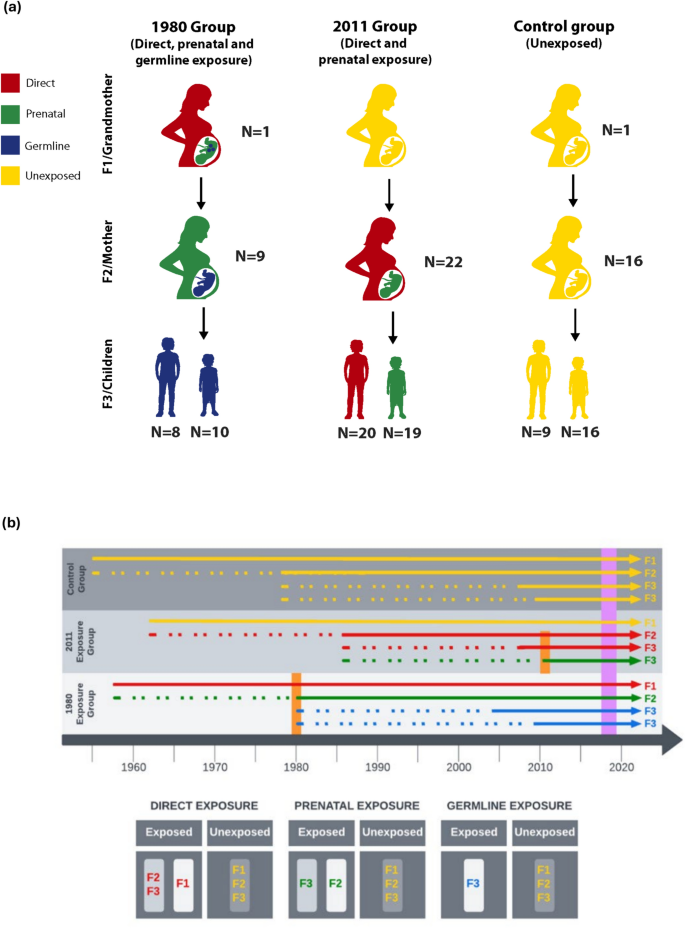
ما از یک طراحی مطالعه سه نسلی برای شناسایی نشانههای DNAm مواجهههای رشدی متضاد با خشونت استفاده کردیم—مواجهه مستقیم یک فرد زنده، مواجهه پیش از تولد یک جنین و مواجهه سلول جنسی. ما گروههای متضادی از خانوادههای سوری سه نسلی را که در حال حاضر در اردن زندگی میکنند، در نظر گرفتیم (شکل ۱). دو گروه اول در معرض خشونتی قرار گرفتند که در طول درگیریهای منطقهای رخ داد: اول، قتل عام شهر حما در سوریه که در سال ۱۹۸۰ آغاز شد، و دوم، قیام سوریه و درگیریهای مسلحانه متعاقب آن که از سال ۲۰۱۱ آغاز شد. در گروه مواجهه ۱۹۸۰، مادربزرگهای مادری قبل از فرار از سوریه باردار بودند، به این معنی که دخترانشان (نسل F2) در معرض پیش از تولد و نوههایشان (نسل F3) در معرض سلول جنسی خشونت جنگ قرار داشتند. در گروه مواجهه ۲۰۱۱، مادران (نسل F2) قبل از فرار از سوریه باردار بودند، به این معنی که جنین در حال رشد در داخل رحم در معرض پیش از تولد قرار داشت، در حالی که کودکان بزرگتر در همان خانواده (نسل F3) به طور مستقیم در معرض درگیری قرار داشتند. در گروه کنترل، مادربزرگها و مادران سوری قبل از سال ۱۹۸۰ در اردن زندگی میکردند. دادههای نظرسنجی و نمونههای سواب دهانی از مادران (نسل F2) و ۱ تا ۲ فرزند (نسل F3) در هر خانواده جمعآوری شد (دو مادربزرگ F1 نیز در این مطالعه گنجانده شدند، یکی در گروه ۱۹۸۰ و دیگری در گروه کنترل). این مطالعه شامل ۱۳۱ شرکتکننده و ۴۸ خانواده بود.
مطالعه انجمنی در مورد کل ژنوم
دادههای DNAm با استفاده از Illumina EPIC BeadChip تولید شدند که بیش از ۸۵۰۰۰۰ سایت CpG را مورد سنجش قرار میدهد. ما یک EWAS انجام دادیم تا موقعیتهای متیله شده متفاوتی (DMPs) را شناسایی کنیم که با هر مواجهه با خشونت مرتبط بودند. از یک روش تحلیلی دو مرحلهای استفاده شد. در مرحله اول، ما سه EWAS جداگانه از آسیبهای ناشی از خشونت را با استفاده از رگرسیون خطی قوی با خطاهای استاندارد قوی انجام دادیم. برای هر دسته از آسیبهای ناشی از خشونت (مستقیم، پیش از تولد یا سلول جنسی)، مقادیر بتا خاص محل به عنوان تابعی از مواجهه با آسیبهای ناشی از خشونت نسبت به کنترلها (بله/خیر) مدلسازی شد، و برای سن در هنگام جمعآوری سواب دهانی، جنسیت و تخمینهای مبتنی بر DNAm از نسبت سلولهای اپیتلیال تنظیم شد. در مرحله دوم، مقادیر بتا خاص محل به عنوان تابعی از همان متغیرهای کمکی با استفاده از معادلات تخمین تعمیمیافته (GEE) مدلسازی شد تا خوشهبندی بر اساس خانواده در نظر گرفته شود. این روش دو مرحلهای از توانایی رگرسیون قوی برای به حداقل رساندن تأثیر نقاط پرت DNAm استفاده کرد، در حالی که GEE وابستگی خانوادگی را که ذاتی طراحی مطالعه بود، در نظر گرفت. مجموعه نهایی DMPs با همپوشانی در DMPs که اصلاح سختگیرانه Bonferroni را برای آزمایشهای متعدد (مقدار p—6.5E-8) در هر دو رگرسیون قوی و EWAS GEE برآورده میکرد، تعیین شد.
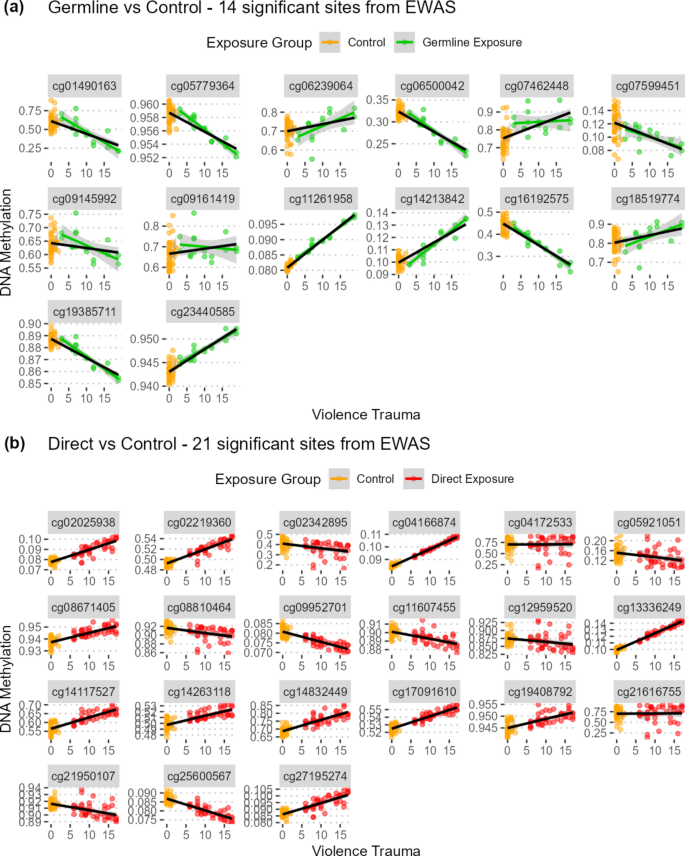
مجموعه نهایی ۳۵ DMP شامل ۱۴ کاوشگر بود که با مواجهه سلول جنسی با خشونت و ۲۱ کاوشگر که با مواجهه مستقیم با خشونت مرتبط بودند (شکل ۲، جدول مکمل S1). هیچ DMP به طور قابل توجهی با مواجهه پیش از تولد با خشونت مرتبط نبود. این اولین گزارش از یک نشانه DNAm سلول جنسی از آسیبهای ناشی از خشونت در انسان با استفاده از یک مطالعه گروهی با یک گروه مقایسه غیرمواجهه است که فرضیه ما مبنی بر یک نشانه اپیژنتیکی بین نسلی خشونت را تأیید میکند. هیچ همپوشانی در DMPs مرتبط با مواجهه سلول جنسی در مقابل مواجهه مستقیم با خشونت وجود نداشت. با این حال، ۳۲ DMP جهت یکسانی را در تفاوتهای DNAm در سراسر هر سه مواجهه نشان دادند، که نشان میدهد این DMPs ممکن است پاسخ اپیژنتیکی یکسانی به خشونت در سراسر مواجههها داشته باشند (شکل ۲).
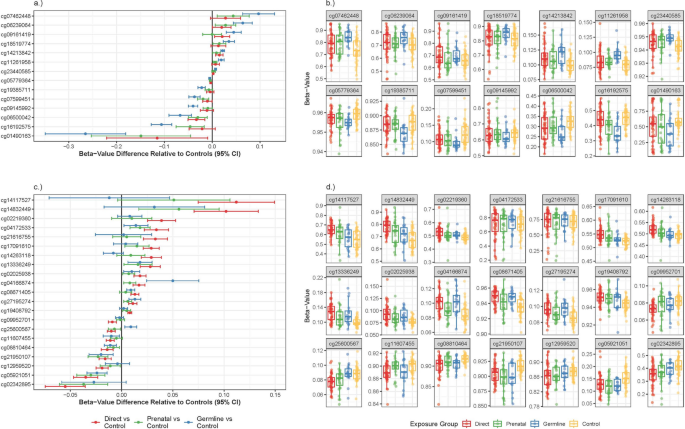
بزرگترین تفاوت در DNAm نسبت به کنترلهای غیرمواجهه در یک DMP مرتبط با سلول جنسی، cg01490163، با DNAm کمتر در بین کسانی که در سلول جنسی در معرض خشونت قرار گرفتهاند (تفاوت: - 0.265، فاصله اطمینان ۹۵٪ (CI) - 0.349، - 0.181) مشاهده شد (شکل ۲). این سایت تقریباً ۳ کیلوبایت بالادست کراتین ۳۶ (KRT36) است که کراتین تولید میکند و نقش بالقوهای در برخی سرطانها دارد و تغییرپذیری بین فردی بارزی در DNAm داشت. در مقایسه با کنترلها، بالاترین DNAm در DMP مرتبط با سلول جنسی، cg07462448 و دو DMP مرتبط با مستقیم، cg14117527 و cg14832449 مشاهده شد. سایت cg07462448 به کاسپاز ۷ (CASP7) اختصاص داده شده است که به خانوادهای از پروتئینها تعلق دارد که نقش محوری در آپوپتوز سلولی ایفا میکنند. سایت cg14117527 به RAB43/ISY1-RAB43 اختصاص داده شده است (RAB43 در مسیرهای انتقال غشایی و هم ولوجی شرکت دارد، و ISY1-RAB43 یک RNA غیرکدکننده است که در یک جهت از RAB43 نشانه میگیرد). سایت cg14832449 به کرومودومین Y-کروموزوم (CDY1) اختصاص داده شده است که به خانواده CDY تعلق دارد که به نظر میرسد در بیان ژن در بیضهها نقش دارد.
استنتاج سن اپیژنتیکی در ارتباط با مواجهه با خشونت در رحم
از آنجایی که با کمبود DMPs برای مواجهه پیش از تولد مواجه شدیم، این ایده را بررسی کردیم که آیا مواجهه با خشونت با تسریع DNAm در میان اعضای گروه مطالعه مرتبط است یا خیر. سن DNAm یک تخمین متمایز DNAm از سن بیولوژیکی است و به عنوان یک شاخص از سلامتی زیستی، مرگومیر و طول عمر در نظر گرفته شده است 40,41. سن اپیژنتیکی شتابگرفته به عنوان تفاوت بین سن DNAm فرد و سن زمانی او تعریف میشود. از میان انواع مختلف ساعت اپیژنتیکی در دسترس، ساعت هورواث بهترین عملکرد را در بازنمایی سن زمانی واقعی در سواب دهانی نشان داده است و برای اندازهگیری شتاب سن اپیژنتیکی 38 در این مطالعه از آن استفاده کردیم. از رگرسیون خطی برای آزمایش ارتباط بین انواع مواجهه و شتاب سن استفاده کردیم. علاوه بر سن در هنگام جمعآوری نمونه سواب دهانی، جنسیت و تخمینهای مبتنی بر DNAm از نسبت سلولهای اپیتلیال، ما همچنین برای نسبتهای سلولهای خونی تنظیم کردیم، زیرا نشان داده شده است که آنها شتاب سن اپیژنتیکی را مخدوش میکنند 42.
در مدل رگرسیون ما که در آن همه شرکتکنندگان گنجانده شده بودند، شتاب سن هورواث به طور قابل توجهی با مواجهه پیش از تولد با خشونت مرتبط بود. شتاب به طور میانگین 3.81 سال بیشتر بود (فاصله اطمینان ۹۵٪ (CI) 0.30، 7.32) در میان کسانی که در معرض رحم قرار گرفته بودند نسبت به کسانی که در معرض رحم قرار نگرفته بودند. شتاب سن هورواث به طور قابل توجهی با مواجهه مستقیم (p؟=?0.982) یا سلول جنسی (p؟=?0.209) مرتبط نبود (شکل 3). این اولین گزارش از ارتباط بین شتاب سن DNAm و مواجهه با خشونت در رحم در انسان است. از آنجایی که نمونه سواب دهانی شامل مخلوطی از سلولهای اپیتلیال و سلولهای خونی است، ما این ایده را بررسی کردیم که آیا نتایج ما به دلیل تغییر در نسبت انواع سلولی به دلیل مواجهه با استرس مرتبط است یا خیر. برای رفع این نگرانی، ما نسبت سلولهای خونی را از DNAm شرکتکنندگان خود با استفاده از روشی مبتنی بر DNAm که به طور گسترده مورد استفاده قرار میگیرد 43 برآورد کردیم و سپس برای آنها در رگرسیون تنظیم کردیم (به «روشها» مراجعه کنید). به طور قابل توجهی، ارتباط بین مواجهه پیش از تولد و شتاب سن DNAm قوی و همچنان از نظر آماری معنیدار باقی ماند (تخمین 4.26، فاصله اطمینان ۹۵٪ (CI) 0.71، 7.81، p؟=?0.019). برای بررسی این موضوع بیشتر، ما این تحلیل را فقط به کودکان محدود کردیم تا اثرات قرار گرفتن در معرض سن را کاهش دهیم. در واقع، ارتباط بین شتاب سن هورواث و قرار گرفتن در معرض رحم به اندازه تقریباً دو برابر تخمین اولیه (تخمین 7.44، فاصله اطمینان ۹۵٪ (CI) 2.30، 12.58، p؟=?0.006) تقویت شد. در نتیجه، این دادهها از اثر مواجهه پیش از تولد بر شتاب سن DNAm در فرزندان، به ویژه در کودکان، پشتیبانی میکنند.
بحث
یافته اصلی مطالعه حاضر نشان میدهد که مواجهه با خشونت در پناهندگان سوری با نشانههای DNAm در محلهای ژنومی خاص و با تسریع سن اپیژنتیکی مرتبط است. ما به طور خاص DNAm متمایز را در میان افرادی که به طور مستقیم، پیش از تولد و سلول جنسی در معرض خشونت قرار گرفتهاند، شناسایی کردیم. به طور جالب توجه، ما شواهدی پیدا نکردیم که نشان دهد سایتهای DNAm تحت تأثیر مواجهه پیش از تولد در EWAS ما هستند، در عوض یک تجزیه و تحلیل جداگانه از شتاب سن اپیژنتیکی وجود یک اثر قابل توجه را آشکار کرد، به طوری که کودکان در گروه مواجهه پیش از تولد در مقایسه با کسانی که در معرض مواجهه قرار نگرفتهاند شتاب سن اپیژنتیکی بالاتری داشتند. با هم، این نتایج از فرضیه ما مبنی بر این که مواجهه با خشونت ممکن است نشانههای اپیژنتیکی بین نسلی ایجاد کند حمایت میکند و پیامدهای مهمی برای درک ما از وراثت آسیب روحی دارد.
ما ۳۵ DMP را شناسایی کردیم که با مواجهه سلول جنسی (۱۴ DMP) و مواجهه مستقیم با خشونت (۲۱ DMP) مرتبط هستند. این نشانههای DNAm با تفاوتهای سطح ژنوم در متیلاسیون DNA خاص محل هنگام مقایسه گروههای مواجهه با خشونت و کنترلها مشخص میشوند. در حالی که ما هیچ همپوشانی بین دو فهرست DMP پیدا نکردیم، ۳۲ DMP در هر دو تجزیه و تحلیل با یکدیگر مطابقت داشتند، که به سمت تفاوت DNAm یکسانی جهتگیری داشتند. برای مثال، در گروه مواجهه سلول جنسی، ما DNAm کمتر را در ناحیه ژنی KRT36 شناسایی کردیم. ژن KRT36 کراتین تولید میکند و نقش بالقوهای در برخی سرطانها دارد و تغییرپذیری بین فردی بارزی در DNAm دارد. کراتینها پروتئینهای ساختاری هستند که در سلولهای اپیتلیال وجود دارند و به عنوان سیتواسکلت در نظر گرفته میشوند که از یکپارچگی ساختاری آنها محافظت میکند و همچنین در تنظیم فرایندهای سلولی متعددی مانند تکثیر سلولی، مهاجرت سلولی و تمایز سلولی شرکت میکنند 44. نقش دقیق KRT36 در بیماریهای انسانی تا حد زیادی ناشناخته باقی مانده است. در مقابل، بالاترین DNAm در DMP مربوط به سلول جنسی، CASP7 و دو DMP مربوط به مواجهه مستقیم—RAB43/ISY1-RAB43 و CDY1 مشاهده شد. ژن CASP7 که به کاسپاز ۷ اختصاص داده شده است، به خانوادهای از پروتئینها تعلق دارد که نقش محوری در آپوپتوز سلولی ایفا میکنند 45. به همین ترتیب، نقش دقیق این ژنها در بیماریهای انسانی ناشناخته باقی مانده است.
یافته ما مبنی بر ارتباط بین مواجهه پیش از تولد با خشونت و شتاب سن اپیژنتیکی با مطالعات قبلی در مورد اثرات نامطلوب مواجهه پیش از تولد بر سلامتی، مرگومیر و طول عمر 46,47 سازگار است. استنتاج سن DNAm از افراد از نظر زمانی بسیار پایدار است، توسط ژنتیک تعیین میشود و در دوران کودکی و نوجوانی به شدت به سن زمانی همبستگی دارد، اما در سنین بالاتر تضعیف میشود 48. شتاب سن DNAm، به جای سن DNAm، به استرس، سبک زندگی ناسالم و قرار گرفتن در معرض تنشها مرتبط است 39,41,49. ما شتاب سن اپیژنتیکی را در شرکتکنندگان در معرض رحم شناسایی کردیم که میتواند گواهی بر این باشد که جنین انسان به اثرات استرسآور قرار گرفتن در معرض رویدادهای روانی-اجتماعی در مقایسه با سایر انواع مواجهه در این مطالعه (مانند سلول جنسی یا مواجهه مستقیم) بسیار حساس است. سن در داخل رحم ممکن است به ویژه به تغییرات اپیژنتیکی حساس باشد زیرا یک دوره تمایز سلولی سریع و تکرار و برنامهریزی مجدد اپیژنتیکی است که در آن استرسها، مانند مواجهه با خشونت، میتوانند مکانهای DNAm خاص ژنوم را تغییر دهند. در این سن، تفاوت بین سن زمانی و بیولوژیکی (به عنوان مثال، شتاب سن اپیژنتیکی) میتواند اثرات پایداری بر رشد و سلامتی بعدی داشته باشد.
برخی از محدودیتهای مطالعه حاضر مستلزم احتیاط در تفسیر است. اول، حجم نمونه ما نسبتاً کوچک بود (۴۸ خانواده، ۱۳۱ شرکتکننده)، اگرچه طرح خانوادگی ما به ما این امکان را میدهد که برخی از واریانسهای غیرقابل محاسبه بین افراد را به دلیل شباهتهای ژنتیکی و محیطی به حداقل برسانیم. دوم، گروه مقایسه ما شامل افراد مقیم اردن قبل از درگیری سوریه بود. اگرچه این گروه به مواجهه با خشونت دسترسی نداشتند، اما احتمالاً تفاوتهای ساختاری در فرصت و ثروت اجتماعی-اقتصادی وجود داشت که نمیتوانیم به طور کامل برای آنها تنظیم کنیم. سوم، یافتههای ما عمدتاً بر اساس DNAm یک بافت منفرد، سواب دهانی است. سوابهای دهانی یک جایگزین کمتر تهاجمی و در دسترس برای خون هستند. با این حال، سوابهای دهانی مخلوطی ناهمگن از سلولهای اپیتلیال و خونی هستند، که ممکن است به دلیل قرار گرفتن در معرض استرس مرتبط با خشونت رخ دهند و نتایج ما را مخدوش کنند. در تلاش برای کاهش این نگرانی، ما سوابهای دهانی را که ما به دست آوردیم برای انواع سلولی تخمینی آنها تنظیم کردیم و یافتههای ما با و بدون تنظیم برای ترکیب سلولی یکسان باقی ماندند، که نشان میدهد ترکیب سلولی تفاوتهای DNAm ما را به طور کامل توضیح نمیدهد. برای تأیید وجود این مکانیسمهای اپیژنتیکی بین نسلی، تحقیقات بیشتر در مورد بافتها در مکانهای آناتومیکی متعدد، از جمله مغز، ضروری است. با وجود این محدودیتها، مطالعه ما قدرتمند است زیرا اولین مطالعه است که از یک طرح سه نسلی با مواجهات متمایز با خشونت برای تجزیه و تحلیل نقش نشانههای DNAm و شتاب سن DNAm در وراثت بین نسلی آسیبهای روانی استفاده میکند.
در نتیجه، این نتایج نشان میدهد که مواجهه با خشونت ممکن است نشانههای اپیژنتیکی بین نسلی ایجاد کند و پیامدهای مهمی برای درک ما از وراثت آسیبهای روحی دارد. این نتایج جدید مسیرهای امیدوارکنندهای را برای آینده پژوهش با تمرکز بر بهبود پیامدهای سلامتی برای افراد و خانوادههایی که از اثرات عمیق و طولانی مدت استرس رنج میبرند، باز میکند.